
Jak ewolucja tłumaczy złożoność życia na Ziemi?
Wprowadzenie
Na pierwszy rzut oka, złożone struktury biologiczne mogą wyglądać, jakby zostały stworzone od razu w obecnej formie, wraz ze wszystkimi, w pełni wykształconymi, współgrającymi komponentami. Nie do pomyślenia wydaje się, że mogłyby one powstać na drodze małych kroków – w wyniku stopniowej ewolucji. W „Czarnej skrzynce Darwina”, Michael Behe wprowadza termin (używany później przez Behe i innych zwolenników Inteligentnego Projektu), który ma odzwierciedlać naturę tej obserwacji – nieredukowalną złożoność. Żadna z części nieredukowalnie złożonego systemu nie spełnia żadnej oczywistej funkcji w oderwaniu od pozostałych elementów.
Behe sugeruje, że poszczególne części nieredukowalnie złożonych struktur biologicznych byłyby bezużyteczne, gdyby nie pojawiły się jednocześnie, oraz że ewolucja nie dysponuje żadnym mechanizmem, który byłby w stanie konstruować tego rodzaju struktury. Dobór naturalny działa wszak metodą małych kroków. Co więcej, nie posiada on zdolności przewidywania. Jeśli jakaś konkretna zmiana ma zostać zachowana, musi ona dostarczyć organizmowi (i jego potomkom) pewne dodatkowe korzyści – choćby były one naprawdę niewielkie. W tym miejscu Behe dokonuje jednak nadmiernego uproszczenia. Teoria ewolucji przewiduje bowiem, że w niewielkich populacjach zmiany neutralne – lub nawet te, które szkodzą organizmowi w małym stopniu – mogą zostać utrwalone. Jednak wciąż, w szerszym kontekście, Behe zdaje się mieć rację. Przyjrzyjmy się zatem, co mówią biolodzy ewolucyjni na temat stopniowej budowy złożonych struktur.
Behe sugeruje, że poszczególne części nieredukowalnie złożonych struktur biologicznych byłyby bezużyteczne, gdyby nie pojawiły się jednocześnie, oraz że ewolucja nie dysponuje żadnym mechanizmem, który byłby w stanie konstruować tego rodzaju struktury. Dobór naturalny działa wszak metodą małych kroków. Co więcej, nie posiada on zdolności przewidywania. Jeśli jakaś konkretna zmiana ma zostać zachowana, musi ona dostarczyć organizmowi (i jego potomkom) pewne dodatkowe korzyści – choćby były one naprawdę niewielkie. W tym miejscu Behe dokonuje jednak nadmiernego uproszczenia. Teoria ewolucji przewiduje bowiem, że w niewielkich populacjach zmiany neutralne – lub nawet te, które szkodzą organizmowi w małym stopniu – mogą zostać utrwalone. Jednak wciąż, w szerszym kontekście, Behe zdaje się mieć rację. Przyjrzyjmy się zatem, co mówią biolodzy ewolucyjni na temat stopniowej budowy złożonych struktur.
Pozornie nieredukowalny system
W swoim podręczniku „Developmental Biology, Eight Edition” Scott Gilbert pokazuje na przykładzie połączonych kości ucha środkowego, jak pozornie nieredukowalne struktury mogą powstać w wyniku stopniowych zmian i doboru naturalnego. Ryby dysponują specjalnym narządem zwanym linią boczną, który rozciąga się wzdłuż ich ciała i pozwala wykrywać wibracje w wodzie. Mają również ucho środkowe, które przydaje się do utrzymywania równowagi i wspomaga linię boczną w wykrywaniu wibracji. Wraz z przeprowadzką niektórych organizmów wodnych na ląd, linia boczna stała się niepotrzebna, gdyż znacznie bardziej przydatne okazało się wykrywanie wibracji powietrza. Kość służąca za podporę czaszki uformowała wówczas strzemiączko. Poza sprawowaniem dotychczasowej funkcji, zaczęła ona również przekazywać fale dźwiękowe – które częściowo odbieramy dzięki szczęce i innym kościom czaszki – do ucha środkowego. Skąd wiemy, że to wciąż ta sama kość? Dowiadujemy się o tym śledząc rozwój embrionalny ryb i gadów. U gadów występuje tylko jedna kość, która przekazuje wibracje powietrza do ucha środkowego – właśnie strzemiączko.
Możemy również prześledzić pochodzenie dwóch pozostałych kostek ucha środkowego – młoteczka i kowadełka – spoglądając na skamieniałości pierwszych ssaków pochodzące sprzed 230 milionów lat. Do tego momentu dwie kości – kość stawowa i kość kwadratowa – służyły jako zawias szczęki. Badacze są jednak przekonani, że spełniały one jeszcze jedną funkcję. Ze względu na ich położenie w bezpośrednim sąsiedztwie strzemiączka, uważa się, że one również uczestniczyły w przekazywaniu fal dźwiękowych.
Możemy również prześledzić pochodzenie dwóch pozostałych kostek ucha środkowego – młoteczka i kowadełka – spoglądając na skamieniałości pierwszych ssaków pochodzące sprzed 230 milionów lat. Do tego momentu dwie kości – kość stawowa i kość kwadratowa – służyły jako zawias szczęki. Badacze są jednak przekonani, że spełniały one jeszcze jedną funkcję. Ze względu na ich położenie w bezpośrednim sąsiedztwie strzemiączka, uważa się, że one również uczestniczyły w przekazywaniu fal dźwiękowych.
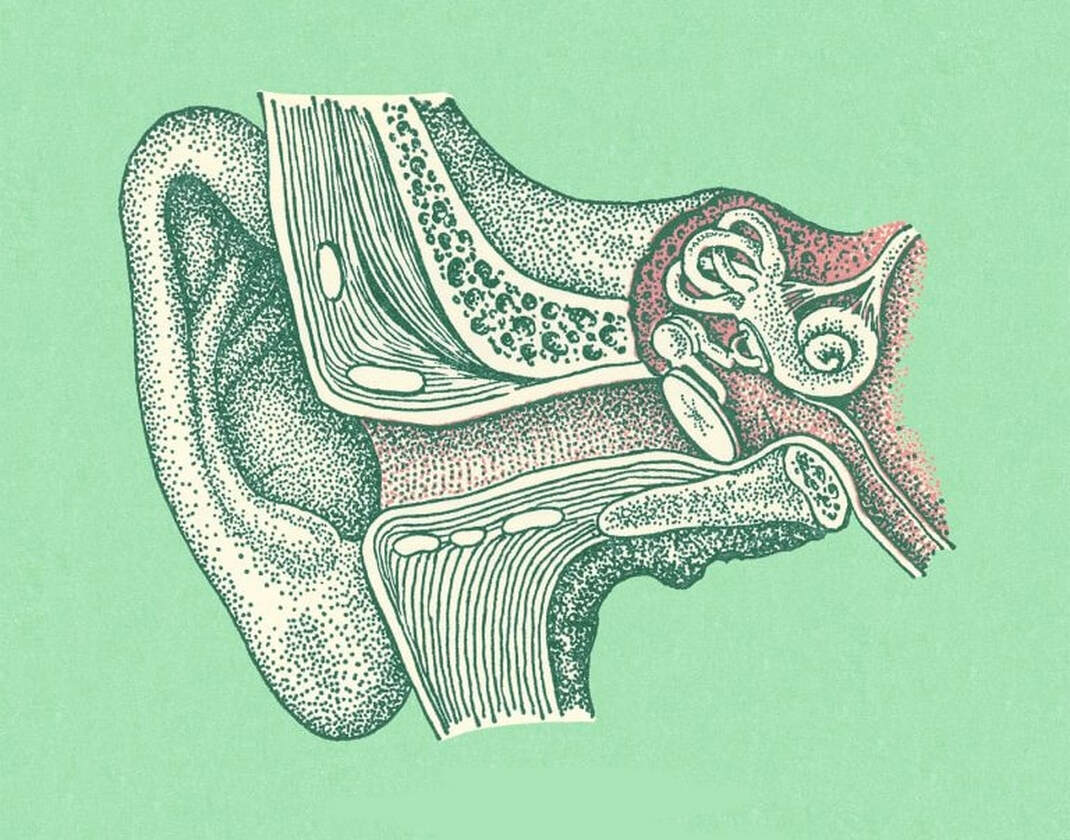
To właśnie w tym momencie nasza opowieść staje się szczególnie interesująca. W czasach pierwszych ssaków pojawiło się kilka gatunków – a być może więcej, paleontolodzy zdają sobie sprawę z niekompletności dostępnego zapisu kopalnego – które dysponowały podwójnym zawiasem szczękowym. Poza parą kości – stawową i kwadratową – również inna para kości – zębowa i łuskowa – spełniały analogiczną funkcję. Dzięki temu te pierwsze, które odpowiadały za transmisję dźwięku, nie musiały już spełniać roli zawiasu. Funkcja ta stała się zbędna, gdyż spełniała ją inna para kości.
Dzięki temu, kości – stawowa i kwadratowa – mogły swobodnie uformować się w młoteczek i kowadełko. Posiadamy zapis tego przejścia, co oznacza, że udokumentowaliśmy powstanie tak zwanej nieredukowalnie złożonej struktury. Części, które początkowo spełniały jedną funkcję, z biegiem czasu stały się użyteczne również z innego powodu. Następnie, pierwsza funkcja została udoskonalona, podczas gdy ta druga okazała się niepotrzebna. Innymi słowy, struktury, które początkowo służyły jednemu celowi, zostały przystosowane do spełniania nowej funkcji. Spoglądając w przeszłość przez pryzmat zapisu kopalnego, możemy ujrzeć świadectwo tej transformacji.
Dzięki temu, kości – stawowa i kwadratowa – mogły swobodnie uformować się w młoteczek i kowadełko. Posiadamy zapis tego przejścia, co oznacza, że udokumentowaliśmy powstanie tak zwanej nieredukowalnie złożonej struktury. Części, które początkowo spełniały jedną funkcję, z biegiem czasu stały się użyteczne również z innego powodu. Następnie, pierwsza funkcja została udoskonalona, podczas gdy ta druga okazała się niepotrzebna. Innymi słowy, struktury, które początkowo służyły jednemu celowi, zostały przystosowane do spełniania nowej funkcji. Spoglądając w przeszłość przez pryzmat zapisu kopalnego, możemy ujrzeć świadectwo tej transformacji.
Wić bakteryjna
W „Czarnej skrzynce Darwina” Behe zwraca uwagę na trzy systemy, które uznaje za nieredukowalnie złożone: wić bakteryjną, mechanizm krzepnięcia krwi i system immunologiczny. Podzespoły tych systemów mają naturę molekularną, co sprawia, że etapy pośrednie są dużo trudniejsze do przebadania. Współgrające cząsteczki nie są utrwalane i nie podlegają analizie historycznej podobnej do tej, którą przeprowadziliśmy w wypadku skamieniałych kości czaszki i ucha środkowego. W swojej książce, Behe sugeruje, że biochemia nie oferuje żadnego wyjaśnienia, w jaki sposób mogłyby powstać mechanizmy złożone z tak wielu współgrających części. Autor stwierdza, że dociekania utknęły w martwym punkcie.
Od publikacji „Czarnej skrzynki Darwina” minęło już ponad 20 lat. Struktury, na których skupił się Behe były w tym czasie starannie badane. Mimo że niemożliwe jest cofnięcie się w czasie, aby krok po kroku odtworzyć to, co się wydarzyło, na przestrzeni lat udało się zgromadzić wiele dowodów popierających wyjaśnienia ewolucyjne. Różnice w budowie tej samej struktury u różnych gatunków pomagają nam zrozumieć, w jaki sposób powstała.
Od publikacji „Czarnej skrzynki Darwina” minęło już ponad 20 lat. Struktury, na których skupił się Behe były w tym czasie starannie badane. Mimo że niemożliwe jest cofnięcie się w czasie, aby krok po kroku odtworzyć to, co się wydarzyło, na przestrzeni lat udało się zgromadzić wiele dowodów popierających wyjaśnienia ewolucyjne. Różnice w budowie tej samej struktury u różnych gatunków pomagają nam zrozumieć, w jaki sposób powstała.
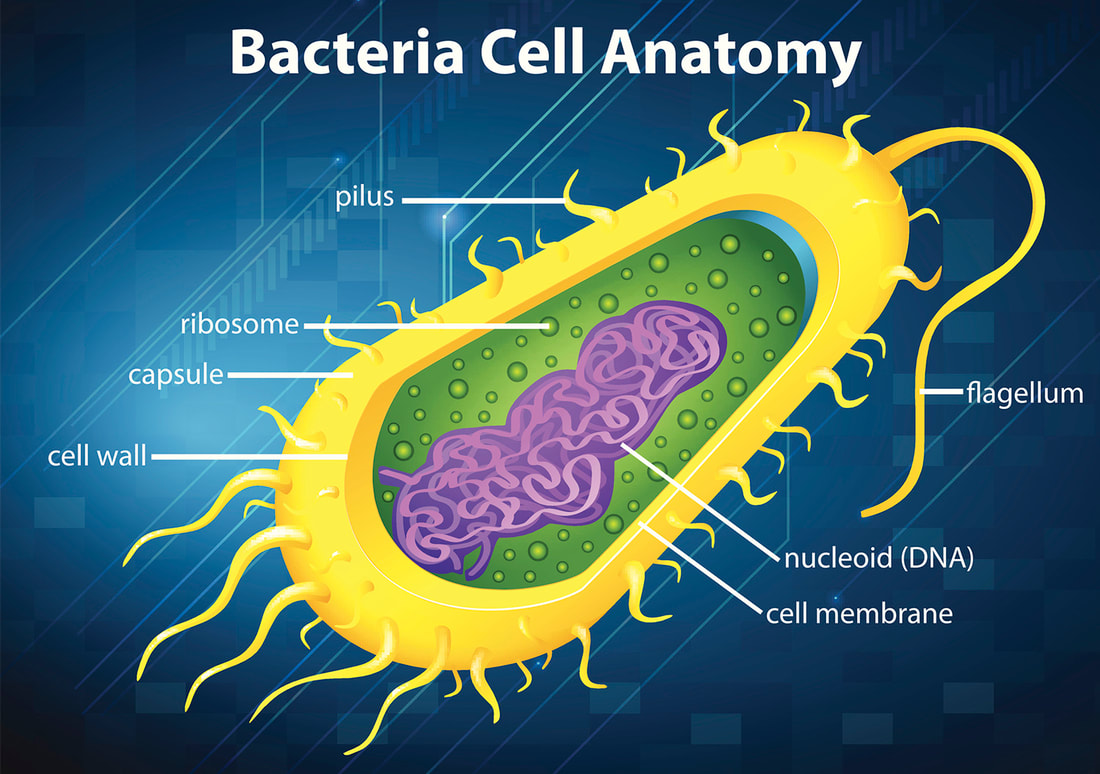
Dla przykładu, istnieje wiele gatunków bakterii wykorzystujących niektóre części swoich wici, aby przekazywać toksyny do komórek nosiciela. Inne gatunki wykorzystują pozostałe części systemu do innych potrzeb. Gatunki z rodzaju Buchnera żyją w zamkniętym środowisku komórek mszyc, w relacji symbiotycznej. Bakterie te nie potrzebują już wici. Jednak każda z komórek posiada setki kopii elementów tworzących bazę dla mechanizmu poruszającego wicią.
Niedawne badanie opublikowane w periodyku „Trends in Microbiology” demonstruje, że struktury te służą jako kanały transportu białek i innych substancji na zewnątrz bakterii, do otaczającego środowiska – komórki mszycy, w której mieszka bakteria. Zatem, o ile faktycznie nie jesteśmy w stanie odtworzyć dokładnego przebiegu wypadków krok po kroku, o tyle możemy zaobserwować w jaki sposób zmienia się konstrukcja mechanizmu i przystosowanie go – w niektórych wypadkach – do pełnienia nowej funkcji. Określenie, którego używa się do opisania sytuacji, w której istniejąca struktura zostaje wykorzystana do roli odmiennej od tej, którą odgrywała wcześniej, to „egzaptacja”. To między innymi dzięki niej możemy obserwować wzrost złożoności organizmów.
Nie jest to obserwacja zupełnie nowa, gdyż już wcześniej obserwowano wzbogacenie istniejących struktur o nowe elementy, aby poprawić ich funkcjonalność lub przystosować do pełnienia nowej funkcji. Skąd biorą się te nowe części? Całkiem niedawno, badacze wykazali, że białko odgrywające kluczową rolę w molekularnym silniku, który wprawia wić w ruch, ma bardzo podobną strukturę do innego białka wykorzystywanego przy transporcie magnezu do wewnątrz i na zewnątrz komórki. Oba białka dysponują fragmentami, które składają się w niemal identyczny sposób. Gdy przyjrzymy się budującym je cegiełkom (aminokwasom), również dostrzeżemy fundamentalne podobieństwa. Przykład ten ilustruje drugi mechanizm powstawania złożoności: kooptację. Części, które niegdyś służyły do realizacji tylko jednego celu przystosowują się do pełnienia dodatkowej funkcji. Czasami przepis na budowę danej części jest kodowany przez dwa identyczne, zduplikowane geny. Gdy taka sytuacja ma miejsce, kooptacja zachodzi szczególnie łatwo. Pierwsza kopia warunkująca powstanie oryginalnej części jest zachowana, podczas gdy druga – stopniowo modyfikowana przez mutacje i dobór naturalny – coraz lepiej przystosowuje się do pełnienia nowej funkcji. To również przykład trzeciej zasady gromadzenia złożoności: adaptacji przez dobór naturalny.
Co jeszcze ciekawsze, owa rzekomo nieredukowalnie złożona wić bakteryjna okazuje się w ogóle nie być tak nieredukowalna. Dla przykładu, u podstawy wici znajduje się białko – ATPaza, która odpowiada za transport podstawowej jednostki strukturalnej – flageliny – przez długi, pusty w środku kanał, co umożliwia przyrost wici na długość. Jak się okazało, transport flageliny możliwy jest również bez wykorzystania ATPazy. Białko, które uważano za nieodzowny element konstrukcyjny wici bakteryjnej wcale nie jest tak niezbędne. Powyższy przykład ilustruje czwarty mechanizm powstawania złożonych struktur: redundancję. W świecie komórek bardzo często istnieje więcej niż jeden sposób na osiągnięcie pożądanego celu. Gdy ewolucja „majstruje” przy skomplikowanych strukturach, w poszczególnych stadiach mogą pojawić nadmiarowe komponenty. Taki nadprogramowy mechanizm może następnie ulegać stopniowym usprawnieniom.
Niedawne badanie opublikowane w periodyku „Trends in Microbiology” demonstruje, że struktury te służą jako kanały transportu białek i innych substancji na zewnątrz bakterii, do otaczającego środowiska – komórki mszycy, w której mieszka bakteria. Zatem, o ile faktycznie nie jesteśmy w stanie odtworzyć dokładnego przebiegu wypadków krok po kroku, o tyle możemy zaobserwować w jaki sposób zmienia się konstrukcja mechanizmu i przystosowanie go – w niektórych wypadkach – do pełnienia nowej funkcji. Określenie, którego używa się do opisania sytuacji, w której istniejąca struktura zostaje wykorzystana do roli odmiennej od tej, którą odgrywała wcześniej, to „egzaptacja”. To między innymi dzięki niej możemy obserwować wzrost złożoności organizmów.
Nie jest to obserwacja zupełnie nowa, gdyż już wcześniej obserwowano wzbogacenie istniejących struktur o nowe elementy, aby poprawić ich funkcjonalność lub przystosować do pełnienia nowej funkcji. Skąd biorą się te nowe części? Całkiem niedawno, badacze wykazali, że białko odgrywające kluczową rolę w molekularnym silniku, który wprawia wić w ruch, ma bardzo podobną strukturę do innego białka wykorzystywanego przy transporcie magnezu do wewnątrz i na zewnątrz komórki. Oba białka dysponują fragmentami, które składają się w niemal identyczny sposób. Gdy przyjrzymy się budującym je cegiełkom (aminokwasom), również dostrzeżemy fundamentalne podobieństwa. Przykład ten ilustruje drugi mechanizm powstawania złożoności: kooptację. Części, które niegdyś służyły do realizacji tylko jednego celu przystosowują się do pełnienia dodatkowej funkcji. Czasami przepis na budowę danej części jest kodowany przez dwa identyczne, zduplikowane geny. Gdy taka sytuacja ma miejsce, kooptacja zachodzi szczególnie łatwo. Pierwsza kopia warunkująca powstanie oryginalnej części jest zachowana, podczas gdy druga – stopniowo modyfikowana przez mutacje i dobór naturalny – coraz lepiej przystosowuje się do pełnienia nowej funkcji. To również przykład trzeciej zasady gromadzenia złożoności: adaptacji przez dobór naturalny.
Co jeszcze ciekawsze, owa rzekomo nieredukowalnie złożona wić bakteryjna okazuje się w ogóle nie być tak nieredukowalna. Dla przykładu, u podstawy wici znajduje się białko – ATPaza, która odpowiada za transport podstawowej jednostki strukturalnej – flageliny – przez długi, pusty w środku kanał, co umożliwia przyrost wici na długość. Jak się okazało, transport flageliny możliwy jest również bez wykorzystania ATPazy. Białko, które uważano za nieodzowny element konstrukcyjny wici bakteryjnej wcale nie jest tak niezbędne. Powyższy przykład ilustruje czwarty mechanizm powstawania złożonych struktur: redundancję. W świecie komórek bardzo często istnieje więcej niż jeden sposób na osiągnięcie pożądanego celu. Gdy ewolucja „majstruje” przy skomplikowanych strukturach, w poszczególnych stadiach mogą pojawić nadmiarowe komponenty. Taki nadprogramowy mechanizm może następnie ulegać stopniowym usprawnieniom.
Oko
Innym systemem, który często służy za przykład nieredukowalnej złożoności, jest oko. Często zadawane pytania brzmią: Jaka korzyść dla organizmu płynie z posiadania częściowo kompletnego oka? Czy istnieje logiczna sekwencja kroków, które mogłyby doprowadzić do stopniowego wykształcenia – na drodze doboru naturalnego – struktury tak wytwornej jak oko orła? Jaki byłby w ogóle punkt wyjścia?
Wszystkie światłoczułe struktury zwierzęce korzystają z tej samej światłoczułej molekuły – retinalu – pochodnej witaminy A. Retinal może zmieniać swój kształt, gdy absorbuje fotony. Wiąże się on również z białkiem – opsyną. Oba te związki chemiczne współpracują przy odbieraniu bodźców świetlnych.
Cały świat organiczny – włączając w to organizmy jednokomórkowe – funkcjonuje w cyklu dobowym. Zatem, wydaje się prawdopodobne, że najprostsza forma systemów światłoczułych powstała przez egzaptację struktur molekularnych wykorzystywanych do wykrywania światła – nie po to, aby organizm mógł się do niego przybliżyć lub oddalić, ale po to, by mógł zresetować swój zegar biologiczny). Również pochodzenie opsyny stanowi przykład podstawowej zasady budowy złożoności – kooptacji. Opsyna jest jednym z wielu receptorów sprzężonych z receptorami G, które spełniały rozmaite funkcje w różnych momentach historii życia. Gdy taki receptor łączy się ze światłoczułą cząsteczką retinalu, komórka staje się wrażliwa na obecność i nieobecność światła. Mimo że nie dysponujemy kopalnymi przykładami form przejściowych, które umożliwiłyby nam prześledzenie rozwoju oka (tak jak było to wypadku kości ucha środkowego), znamy niezliczone przykłady światłoczułych struktur w świecie współczesnych zwierząt. Pozwalają nam one odtworzyć prawdopodobny przebieg budowy oka na drodze stopniowego procesu kierowanego przez dobór naturalny. (Więcej na temat potencjalnych form przejściowych, które występują w świecie zwierząt, możesz przeczytać w fantastycznej pracy Ryana Gregory’ego).
Wszystkie światłoczułe struktury zwierzęce korzystają z tej samej światłoczułej molekuły – retinalu – pochodnej witaminy A. Retinal może zmieniać swój kształt, gdy absorbuje fotony. Wiąże się on również z białkiem – opsyną. Oba te związki chemiczne współpracują przy odbieraniu bodźców świetlnych.
Cały świat organiczny – włączając w to organizmy jednokomórkowe – funkcjonuje w cyklu dobowym. Zatem, wydaje się prawdopodobne, że najprostsza forma systemów światłoczułych powstała przez egzaptację struktur molekularnych wykorzystywanych do wykrywania światła – nie po to, aby organizm mógł się do niego przybliżyć lub oddalić, ale po to, by mógł zresetować swój zegar biologiczny). Również pochodzenie opsyny stanowi przykład podstawowej zasady budowy złożoności – kooptacji. Opsyna jest jednym z wielu receptorów sprzężonych z receptorami G, które spełniały rozmaite funkcje w różnych momentach historii życia. Gdy taki receptor łączy się ze światłoczułą cząsteczką retinalu, komórka staje się wrażliwa na obecność i nieobecność światła. Mimo że nie dysponujemy kopalnymi przykładami form przejściowych, które umożliwiłyby nam prześledzenie rozwoju oka (tak jak było to wypadku kości ucha środkowego), znamy niezliczone przykłady światłoczułych struktur w świecie współczesnych zwierząt. Pozwalają nam one odtworzyć prawdopodobny przebieg budowy oka na drodze stopniowego procesu kierowanego przez dobór naturalny. (Więcej na temat potencjalnych form przejściowych, które występują w świecie zwierząt, możesz przeczytać w fantastycznej pracy Ryana Gregory’ego).
Jeśli zdecydujesz się na szczegółową analizę historii rozwoju oka, zwróć szczególną uwagę na przykłady egzaptacji, kooptacji, stopniowej adaptacji i redundancji. Istnieją dwa sposoby na wyostrzenie obrazu padającego na światłoczułe komórki w tylnej części oka. Jednym ze sposobów jest wykształcenie niezależnej soczewki. Drugim sposobem jest użycie soczewki schowanej za przezroczystą rogówką. Soczewka ta może składać się z przezroczystych, krystalicznych cząstek białka, które są uformowane w taki sposób, że pozwalają wyostrzyć obraz. Istnieje wiele białek, które podlegają krystalizacji i mogą zostać wykorzystane do stworzenia użytecznej soczewki. Okazuje się, że w zależności od obranej ścieżki ewolucyjnej, organizmy wykorzystują do tego celu różne białka – między innymi enzymy takie jak: dehydrogenaza alkoholowa (enzym rozkładający etanol), S-transferaza glutationu czy białka opiekuńcze. Stanowi to prosty przykład na współgranie kooptacji i redundancji na drodze do budowy skomplikowanej struktury, takiej jak oko.
Dwie trzecie typów zwierząt (typ to jedna z podstawowych kategorii systematycznych) dysponuje jakiegoś rodzaju narządem światłoczułym. Mimo że wszystkie te struktury wykorzystują retinal i opsyny, istnieją inne różnice strukturalne, które łatwo wyjaśnić, analizując historię ewolucji. W 2003 roku Simon Conway-Morris opublikował książkę „Life’s Solution”, gdzie dokumentuje co najmniej siedem ścieżek powstania oka, które niezależnie doprowadziły do bardzo podobnego efektu końcowego. Dla przykładu, oczy kałamarnicy działają zadziwiająco podobnie jak oczy ssaków. Jednak sposób, w jaki formują się obie struktury we wczesnych etapach rozwoju organizmu różni się znacznie. Różnice strukturalne są uwarunkowane przez to, jak konstruowane są poszczególne części ciała podczas rozwoju embrionalnego. W budowie oczu odnajdziemy również wskazówki, które pozwalają stwierdzić, że niektóre etapy ich konstrukcji były mocno prowizoryczne. Oczy nie są perfekcyjne. Mają plamki ślepe, są podatne na odwarstwienie siatkówki, jaskrę czy zwyrodnienie plamki żółtej. Wszystkie te nieprzyjemne przypadłości znajdują uzasadnienie w historii rozwoju oka.
Mimo że nie dysponujemy skamieniałymi przykładami form pośrednich (tak jak w wypadku kości ucha środkowego), posiadamy szeroką gamę zwierzęcych oczu o różnej budowie, z których wszystkie mogą zdawać się być nieredukowalnie złożone. Okazuje się jednak, że one również zostały skonstruowane na drodze procesów takich jak: egzaptacja, kooptacja, stopniowa adaptacja i redundancja. W zasadzie, współcześnie obserwowane struktury również można uznać za formy przejściowe. Wszystko zmienia się, gdy zostaje poddane próbie niezwykle długiego czasu, stanowiąc spuściznę stworzenia na drodze doboru naturalnego.
© BioLogos Foundation | Zdjęcie na początku: bakterie probiotyczne Lactobacillus | Wersja artykułu: 19 stycznia 2019
Dwie trzecie typów zwierząt (typ to jedna z podstawowych kategorii systematycznych) dysponuje jakiegoś rodzaju narządem światłoczułym. Mimo że wszystkie te struktury wykorzystują retinal i opsyny, istnieją inne różnice strukturalne, które łatwo wyjaśnić, analizując historię ewolucji. W 2003 roku Simon Conway-Morris opublikował książkę „Life’s Solution”, gdzie dokumentuje co najmniej siedem ścieżek powstania oka, które niezależnie doprowadziły do bardzo podobnego efektu końcowego. Dla przykładu, oczy kałamarnicy działają zadziwiająco podobnie jak oczy ssaków. Jednak sposób, w jaki formują się obie struktury we wczesnych etapach rozwoju organizmu różni się znacznie. Różnice strukturalne są uwarunkowane przez to, jak konstruowane są poszczególne części ciała podczas rozwoju embrionalnego. W budowie oczu odnajdziemy również wskazówki, które pozwalają stwierdzić, że niektóre etapy ich konstrukcji były mocno prowizoryczne. Oczy nie są perfekcyjne. Mają plamki ślepe, są podatne na odwarstwienie siatkówki, jaskrę czy zwyrodnienie plamki żółtej. Wszystkie te nieprzyjemne przypadłości znajdują uzasadnienie w historii rozwoju oka.
Mimo że nie dysponujemy skamieniałymi przykładami form pośrednich (tak jak w wypadku kości ucha środkowego), posiadamy szeroką gamę zwierzęcych oczu o różnej budowie, z których wszystkie mogą zdawać się być nieredukowalnie złożone. Okazuje się jednak, że one również zostały skonstruowane na drodze procesów takich jak: egzaptacja, kooptacja, stopniowa adaptacja i redundancja. W zasadzie, współcześnie obserwowane struktury również można uznać za formy przejściowe. Wszystko zmienia się, gdy zostaje poddane próbie niezwykle długiego czasu, stanowiąc spuściznę stworzenia na drodze doboru naturalnego.
© BioLogos Foundation | Zdjęcie na początku: bakterie probiotyczne Lactobacillus | Wersja artykułu: 19 stycznia 2019